Metallopeptide Project
Background
Living systems have elaborated processes to prevent the toxic side effects of metals while enabling them to employ essential metals in necessary biological function. Heavy metals such as Hg, As, Cd and Pb are purely toxic. Manifestations of these toxicities in humans include symptoms such as nausea, vomiting, headaches and confusion. Prolonged exposure can result in more serious problems such as respiratory damage, skin lesions, a variety of neurological disorders including brain damage, and death. Recent studies have shown that certain bacteria have developed mechanisms for the detoxification of heavy metals. The expression of the detoxification proteins iso-allosterically controlled by a class of heavy metal sensing proteins cease to inhibit expression of the detoxification genes in the presence of the heavy metals.
Goals of our Project
- To understand metalloprotein structure and folding using simple peptidic constructs.
- To develop peptidic systems that can efficiently and selectively sequester heavy metal ions from aqueous solutions and
- To understand the thermodynamics and kinetics of metal binding to these designed peptides.
- To understand the structural aspects of these designed peptides.
- To develop peptide systems that provide a groundwork for understanding metalloregulatory proteins and metallochaperones
Applications and Current Work
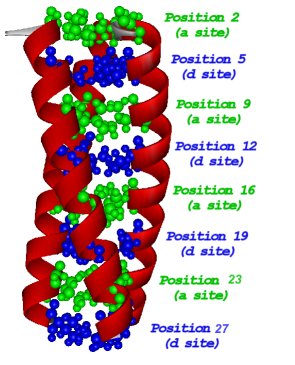
Protein folding is a subject that has attracted the attention of scientists from various disciplines. The protein folding problem can be divided into two categories: the first is the attempt to predict the three dimensional structure of a protein in its native state from a knowledge of the primary sequence and the second would be to understand the relationship between the protein sequence and its mechanism of folding. De Novo design of proteins is a useful technique for understanding the factors that influence metalloprotein structure and stability. Through much of the 1990's we designed and characterized a series of -helical coiled coil peptides. The parent peptide TRI has the amino acid sequence G(LKALEEK)4G.
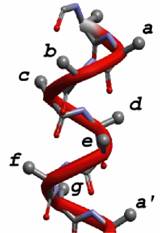
The peptide was designed using the repeated heptad approach. Hydrophobic residues like Leucine(L)were placed in the a and d positions. Hydrophilic residues like Lysine(K) and Glutamic Acid(E) capable of forming salt bridges were placed at postions e and g. Helix inducing residue like Alanine(A) was placed at position c.

This design leads to a pH dependent structure. Protonation of the Glutamate residues at pH values above 7 lead to salt bridges that result in a three helical aggregate. At low pH the system is a two stranded coiled coil.
The hydrophobic residues are arranged in layers. Substitution of Leucine residues in a layer usually with cysteine leads to a soft metal binding site. The orientation of the Cysteine in the core is dependent on the type of site ("a" or "d") that has been modified.
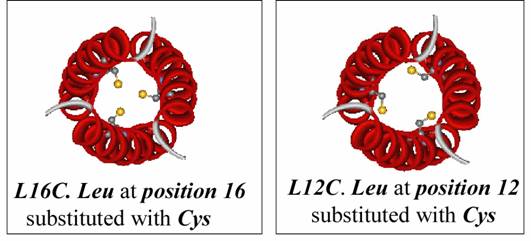
In all of the studies, the chosen metals do not bind to TRI and only form stable complexes with cysteine substituted peptides. Besides TRI, shorter peptides like Baby TRI, Baby L9C, Mini Tri, Mini L9C having 4,4,3 and 3 heptads have also been synthesized and used in our research. Similarly longer peptide like Grand TRI and Grand L9C having one heptad more than TRI have been synthesized and characterized.
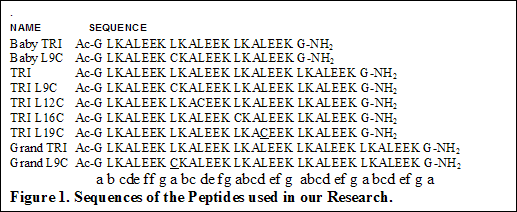
Metal Binding to the Peptides
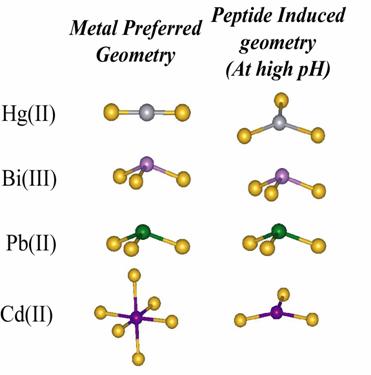
In many cases the geometrical preference of the peptide and the metal are different.The peptide can either form a dimer or a trimer while the metal geometry varies according to the metal prefrence, from two [Hg(II)], three [Bi(III) and Pb(II)], to even higher [Cd(II)]. Our interest is to investigate impact on protein structure and folding that results from the interplay between the metal and the peptide geometry ie. preferences. Investigation of the heavy metal binding is possible with the use of various techniques including UV, CD, EXAFS, PAC and NMR spectroscopy.
Binding of As(III) to the peptides
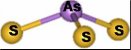
- Arsenic(III) is found primarily with trigonal-pyramidal coordination geometry, with expansion to tetrahedral geometry found in some cases.
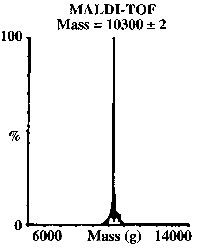
- The addition of excess NaAsO2 to a solution of TRI L16C gave As(TRI L16C)3, which was purified on a reverse phase C18 column in the presence of 0.1% trifloroacetic acid. The figure on the left shows that this complex was verified by mass spectrometry
- These results indicated that As(III) stabilized the three-stranded coiled coil form of the peptide at high pH, where the peptide prefers a three-stranded coiled coil structure. The addition of NaAsO2 to TRI L16C at pH 5.5 also stabilized the trigonal form of the peptide at low pH. This result suggested that formation of As(TRI L16C)3 is not only kinetically but also thermodynamically favored at low pH, a condition where the peptide prefers to be a two-stranded coiled coil.
- Farrer, B.T.; McClure, C.; Penner-Hahn, J.E.; Pecoraro, V.L. "Arsenic(III)-Thiol Interactions Stabilize Three-Helix Bundles in Aqueous Solution"; Inorg. Chem., 2000, 39, 5422-5423.
Binding of Hg(II) to the peptides

Binding of Hg(II) to TRIL16C
- The addition of two equivalents of either TRI L16C to a solution of HgCl2 at high or low pH resulted in the formation of two-stranded coiled coils with Hg(II) bound linearly to two sulfur atoms.
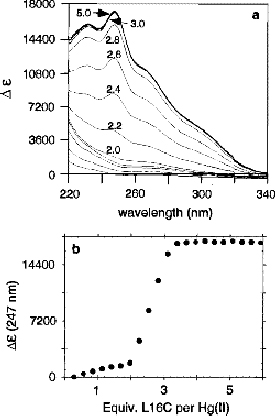
- At pH 4.5, the addition of excess peptide has no effect on the Hg(II)(TRI LC)2 aggregation state. In contrast, the addition of one additional equivalent of TRI L16C at pH 8.5 causes dramatic changes in the metallopeptide. As shown with the difference UV spectra on the right, a strong ligand to metal charge transfer transition develops upon addition of the third peptide equivalent
Farrer, B. T.; Pecoraro, V.L. "Thermodynamic Model for the Stabilization of Trigonal Hg(II) in a Designed Three Helix Bundle," Biochemistry, 2001, 40, 14696-14705.
Binding of Hg(II) to TRIL12C

- Our initial studies of Hg(II) binding by TRI L12C suggested that this d substituted peptide would not bind Hg(II) in a trigonal geometry. This observation suggested that the cysteine conformers in a vs. d positions of the heptad bound Hg differently. We have now investigated this system more fully and realize that the differential behavior lies in the pKa of the cysteine residues in these mercury complexes.
- Figure on the left shows the pH titration curves for Hg(II) with three equivalents of TRI L16C or TRI L12C. There is a marked shift in this titration curve towards higher pH when TRI L12C binding of Hg(II) is monitored.
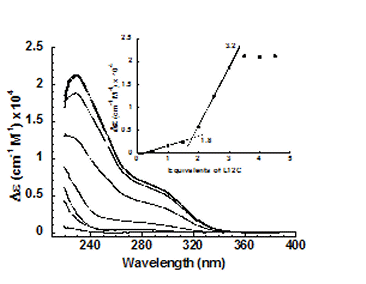
- Difference titrations performed at pH 9.4, showed that the species formed was consistent with Hg(TRI L12C)3 after the addition of the third equivalent of peptide.
- These studies demostrate that the ability to bind a metal within a hydrophobic interior of a helix is critically dependent on the location of the cysteine in the protein sequence.
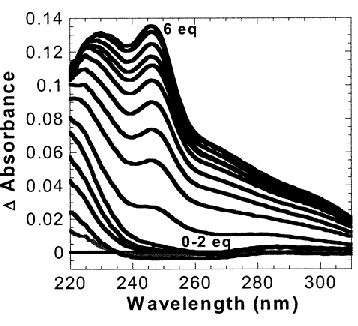
Hg Binding to BabyL9C
- The shorter peptide BabyL9C not only gave the smaller formation constant, but also showed less than 20% helicity at 10M and 5M concentrations in the absence of denaturant, indicating a weakly associated or unassociated coiled coil.
- Interestingly, Baby L9C was shown to stabilize trigonal thiolato Hg(II) within the interior of a three-stranded coiled coil as observed by the characteristic LMCT bands in the UV spectrum.
- The proposed model for this binding involved the initial formation of the two-stranded coiled coil, the addition of the third peptide to form the three-stranded coiled coil and the subsequent deprotonation of a cysteine thiol to generate the Hg(II)(SR)3-. The new model was termed the "Stepwise Aggregation-Deprotonation (StepAD) Model,"
The Kinetic Model
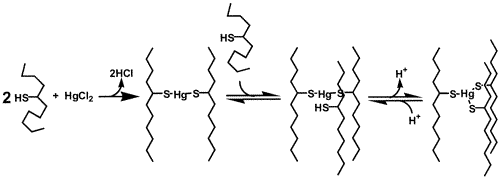
- The StepAD model provided not only a robust explanation for the encapsulation of Hg(II) within the interior of coiled coils, but also provided a basis for a proposed kinetic mechanism for the encapsulation of Hg(II) by unstructured peptides such as Baby L9C (Figure on the left). The proposed kinetic mechanism involves the initial rapid collapse of two peptide chains and a Hg(II) ion to form a two-stranded coiled coiled with linear thiolato Hg(II). The second step, addition of the third peptide to form a three-stranded coiled coil with linear thiolato Hg(II) encapsulated, is rate determining. The third step is a rapid deprotonation yielding the ultimate structure.
Farrer, Brian T.; Pecoraro, Vincent L. "Hg(II) binding to a weaklyassociated coiled coil nucleates an encoded metalloprotein fold: A Kinetic Analysis" Proc. Natl. Acad. Sci., 2003 , 3760-3765.
The general features of the folding mechanism for Hg(Baby L9C)3-, initial collapse of the peptide backbone yielding a structure with a metal in its preferred coordination environment followed by a change in the backbone structure resulting in the final structure of the peptide, resemble mechanisms of natural metalloprotein folding that control the coordination environment around a metal.
Binding of Cd(II) to the peptides

The pH dependent binding of Cd(II) to TRI L12C and TRI L16C, shown in the Figure on the right, was similar to that observed for Hg(II); however, there are two important differences. First, the titration curves with Cd(II) are shifted to lower pH for both TRI L12C and TRI L16C. Note, however, that TRI L16C still contains a slightly more acidic cysteine residue. Second, Cd(II) binds to neither TRI L12C nor TRI L16C at low pH, whereas Hg(II) forms a linear adduct in a two-stranded coiled coil.
Matzapetakis, M.; Farrer, B. T.; Weng, T.-C.; Hemmingsen, L.; Penner-Hahn, J. E.; Pecoraro, V. L.; " Comparison of the Binding of Cadmium(II), Mercury(II), and Arsenic(III) to the de Novo Designed Peptides TRI L12C and TRI L16C" J. Am. Chem. Soc., 2002, 124(27) 8042-8054
Structural Characterization
In the sections above, we described a system that has led to significant new understanding of both heavy metal biochemistry and metal complexation by de novo designed proteins. The following section presents our strategy to gain further insight into both of these important fields. Our approach is unique in that we can generate small proteins that can answer fundamental questions dealing with heavy metal biochemistry and metalloprotein structure and dynamics. We feel it is essential to the next phase of our work to begin correlating structure with the kinetic, thermodynamic and spectroscopic properties that we have uncovered. Also, we believe we know enough from our preliminary work to begin modifying the protein structures to either increase metal loading in a peptide or develop sites for small molecule binding sites to the metal. Furthermore, we feel that it is necessary to take what we have learned about untethered systems and apply these principles to helical bundles.
In this context, we have research undergoing in our lab where the techniques of NMR (COSY, NOESY, TOCSY) and X-Ray Crystallography are being applied to obtain structures of both apo as well as metallated peptides.

An NMR structure of the (TRIL12C)3